Summary
This strain contains 11 protein-level mutations:
- 3 mutations in Spike protein: E484K, D614G, V1176F
- 4 mutations in N protein: A119S, 203-204 RG->KR, M234I, G238C
- 1 mutation in NS9c protein: G50N
- 1 mutation in NSP5 protein: L205V
- 1 mutation in NSP7 protein: L71F
- 1 mutation in NSP12 protein: P323L
We identified all possible linear viral peptides affected by these mutations. Whenever it was possible, we matched the reference peptide with the mutated one. For example, D -> L mutation transformed SDNGPQNQR to SLNGPQNQR. Cases when it was not meaningful included deletions and insertions at the flanks of the peptide, e.g., HV deletion in NVTWFHAIHV peptide.
Then, we predicted binding affinities between the selected peptides and frequent HLA alleles. Predictions were made with NetMHCpan-4.1 and NetMHCIIpan-4.0. The binding affinities were classifies into three groups:
- Tight binding (IC50 affinity ≤ 50 nM)
- Moderate binding (50 nM < IC50 affinity ≤ 500 nM)
- Weak/no binding (IC50 affinity > 500 nM)
Here we report HLA-peptide interactions whose affinity was altered by at least two folds. Note that mutations with empty set of altered interactions are not showed.
The total number of HLA-peptide interactions affected by the mutations
For each allele we calculated the number of peptides with increased and decreased affinitiy (two or more times). The absolute number of such peptides is showed on the left side of the figure (blue bars — weaker binding, orange bars — stronger).
While absolute numbers are of interest, they do not reflect the initial number of tighly binding peptides for the specific allele. Roughly speaking, if an allele had high number of tight binders, then vanished affinity of several peptides will not affect much the integral ability of peptide presentation. To the contrary, low number of vanished peptides could be important if the particular allele initially had narrow epitope repertoire. To account for this effect, we normalized the absolute numbers of new/vanished tight binders (IC50 affinity ≤ 50 nM) by the total number of tight binders for each allele. To avoid possible divisions by zero, we used + 1 regularization term in all denominators. For example, if some allele had no tight binders in the reference genome, and three new tight binders appeared as a result of the mutation, the relative increase will be 300%. The results are showed in the right part of the figure. Protein-specific plots are listed below.
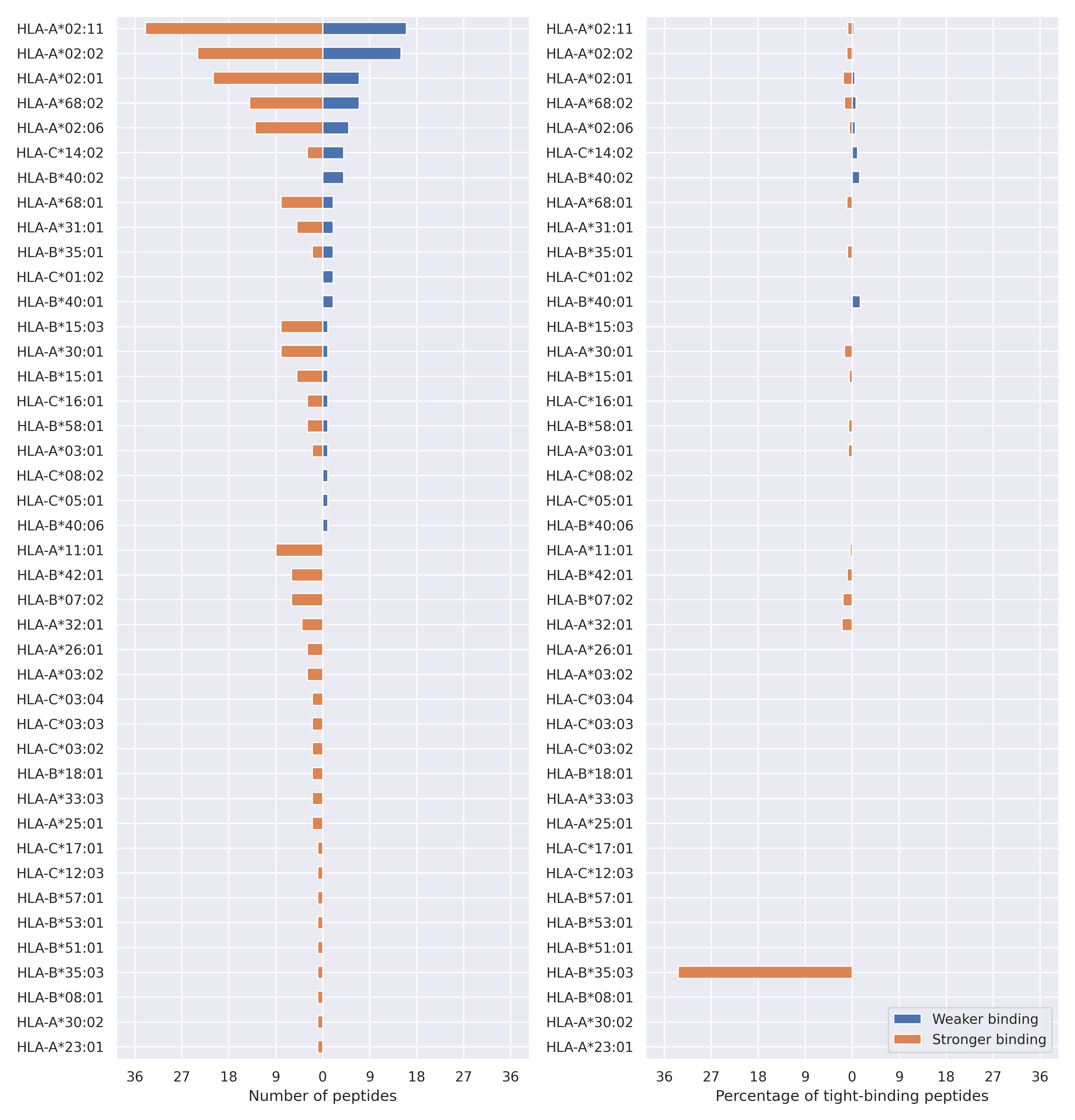{kind=link}

Spike protein

E484K
Reference SNLKPFERDISTEIYQAGSTPCNGVEGFNCYFPLQSYGFQPTNGVGYQPYR
||||||||||||||||||||||||| |||||||||||||||||||||||||
Mutated SNLKPFERDISTEIYQAGSTPCNGVKGFNCYFPLQSYGFQPTNGVGYQPYR
| Allele | Reference peptide | Mutated peptide | Reference affinity (IC50, nM) | Mutated affinity (IC50, nM) |
|---|---|---|---|---|
| HLA-B*40:02 | VEGFNCYFPL | VKGFNCYFPL | 31 | 7011 |
| HLA-B*40:01 | VEGFNCYFPL | VKGFNCYFPL | 76 | 15210 |
| HLA-A*11:01 | GSTPCNGVE | GSTPCNGVK | 32767 | 444 |
| HLA-A*68:02 | EGFNCYFPL | KGFNCYFPL | 117 | 3877 |
| HLA-A*30:01 | EGFNCYFPL | KGFNCYFPL | 5413 | 164 |
| HLA-A*32:01 | EGFNCYFPL | KGFNCYFPL | 6548 | 210 |
| HLA-A*30:02 | EGFNCYFPLQSY | KGFNCYFPLQSY | 1806 | 72 |
| HLA-B*15:01 | GVEGFNCYF | GVKGFNCYF | 3125 | 132 |
| HLA-A*31:01 | EGFNCYFPL | KGFNCYFPL | 4227 | 226 |
| HLA-B*15:03 | EGFNCYFPL | KGFNCYFPL | 2980 | 212 |
| HLA-A*02:11 | EGFNCYFPL | KGFNCYFPL | 2508 | 259 |
| HLA-B*15:03 | GVEGFNCYF | GVKGFNCYF | 1657 | 199 |
| HLA-A*02:06 | EGFNCYFPL | KGFNCYFPL | 1266 | 169 |
| HLA-B*35:01 | TPCNGVEGF | TPCNGVKGF | 319 | 1754 |
| HLA-B*15:03 | VEGFNCYFPLQSY | VKGFNCYFPLQSY | 1081 | 208 |
| HLA-B*35:01 | NGVEGFNCY | NGVKGFNCY | 346 | 1439 |
| HLA-C*16:01 | EGFNCYFPL | KGFNCYFPL | 1861 | 498 |
| HLA-B*15:03 | VEGFNCYFPL | VKGFNCYFPL | 943 | 280 |
D614G
Reference PCSFGGVSVITPGTNTSNQVAVLYQDVNCTEVPVAIHADQLTPTWRVYSTG
||||||||||||||||||||||||| |||||||||||||||||||||||||
Mutated PCSFGGVSVITPGTNTSNQVAVLYQGVNCTEVPVAIHADQLTPTWRVYSTG
| Allele | Reference peptide | Mutated peptide | Reference affinity (IC50, nM) | Mutated affinity (IC50, nM) |
|---|---|---|---|---|
| HLA-C*05:01 | YQDVNCTEV | YQGVNCTEV | 82 | 8597 |
| HLA-C*08:02 | YQDVNCTEV | YQGVNCTEV | 89 | 8066 |
| HLA-A*02:11 | DVNCTEVPV | GVNCTEVPV | 2140 | 72 |
| HLA-A*02:02 | DVNCTEVPV | GVNCTEVPV | 4099 | 213 |
| HLA-A*68:02 | DVNCTEVPV | GVNCTEVPV | 36 | 659 |
| HLA-A*02:06 | DVNCTEVPV | GVNCTEVPV | 3807 | 208 |
| HLA-A*68:02 | QVAVLYQDV | QVAVLYQGV | 183 | 13 |
| HLA-A*68:02 | NQVAVLYQDV | NQVAVLYQGV | 650 | 52 |
| HLA-A*02:06 | QVAVLYQDV | QVAVLYQGV | 1312 | 148 |
| HLA-A*02:11 | QVAVLYQDV | QVAVLYQGV | 2169 | 268 |
| HLA-A*68:02 | NTSNQVAVLYQDV | NTSNQVAVLYQGV | 1858 | 231 |
| HLA-A*68:02 | TSNQVAVLYQDV | TSNQVAVLYQGV | 3281 | 457 |
| HLA-A*02:02 | QVAVLYQDV | QVAVLYQGV | 1107 | 188 |
| HLA-A*02:01 | YQDVNCTEV | YQGVNCTEV | 75 | 315 |
| HLA-A*02:06 | YQDVNCTEV | YQGVNCTEV | 17 | 57 |
| HLA-A*02:11 | VLYQDVNCT | VLYQGVNCT | 224 | 74 |
| HLA-A*02:11 | YQDVNCTEV | YQGVNCTEV | 31 | 93 |
| HLA-A*02:11 | VLYQDVNCTEVP | VLYQGVNCTEVP | 1000 | 402 |
| HLA-A*02:02 | NQVAVLYQDV | NQVAVLYQGV | 269 | 116 |
| HLA-A*02:11 | AVLYQDVNCTEV | AVLYQGVNCTEV | 500 | 238 |
| HLA-A*02:01 | NQVAVLYQDV | NQVAVLYQGV | 558 | 267 |
| HLA-A*02:06 | NQVAVLYQDV | NQVAVLYQGV | 36 | 18 |
V1176F
Reference ELDKYFKNHTSPDVDLGDISGINASVVNIQKEIDRLNEVAKNLNESLIDLQ
||||||||||||||||||||||||| |||||||||||||||||||||||||
Mutated ELDKYFKNHTSPDVDLGDISGINASFVNIQKEIDRLNEVAKNLNESLIDLQ
| Allele | Reference peptide | Mutated peptide | Reference affinity (IC50, nM) | Mutated affinity (IC50, nM) |
|---|---|---|---|---|
| HLA-A*68:02 | DISGINASV | DISGINASF | 10 | 1289 |
| HLA-A*02:11 | GDISGINASV | GDISGINASF | 238 | 15161 |
| HLA-C*14:02 | SVVNIQKEI | SFVNIQKEI | 6508 | 124 |
| HLA-A*02:02 | GDISGINASV | GDISGINASF | 71 | 3625 |
| HLA-A*02:11 | LGDISGINASV | LGDISGINASF | 446 | 19525 |
| HLA-A*68:02 | DLGDISGINASV | DLGDISGINASF | 362 | 15753 |
| HLA-B*15:01 | GDISGINASV | GDISGINASF | 15806 | 370 |
| HLA-A*02:02 | LGDISGINASV | LGDISGINASF | 208 | 8728 |
| HLA-B*35:01 | DISGINASV | DISGINASF | 16065 | 447 |
| HLA-A*68:02 | GDISGINASV | GDISGINASF | 453 | 15954 |
| HLA-A*02:02 | VDLGDISGINASV | VDLGDISGINASF | 413 | 11912 |
| HLA-A*68:01 | SVVNIQKEIDR | SFVNIQKEIDR | 288 | 5363 |
| HLA-A*02:02 | VVNIQKEIDRLNEV | FVNIQKEIDRLNEV | 2908 | 166 |
| HLA-A*25:01 | DISGINASV | DISGINASF | 3057 | 204 |
| HLA-A*02:06 | VVNIQKEIDRLNEV | FVNIQKEIDRLNEV | 871 | 59 |
| HLA-A*26:01 | DISGINASV | DISGINASF | 1865 | 129 |
| HLA-A*02:11 | VVNIQKEIDRLNEV | FVNIQKEIDRLNEV | 2044 | 152 |
| HLA-A*68:01 | VVNIQKEIDR | FVNIQKEIDR | 1917 | 242 |
| HLA-A*02:06 | GINASVVNI | GINASFVNI | 1251 | 237 |
| HLA-A*68:01 | SGINASVVNIQK | SGINASFVNIQK | 913 | 194 |
| HLA-A*68:01 | NASVVNIQKEIDR | NASFVNIQKEIDR | 127 | 30 |
| HLA-A*68:01 | INASVVNIQK | INASFVNIQK | 249 | 62 |
| HLA-A*11:01 | ASVVNIQK | ASFVNIQK | 1597 | 479 |
| HLA-A*68:02 | DISGINASVV | DISGINASFV | 59 | 19 |
| HLA-A*03:02 | NASVVNIQK | NASFVNIQK | 949 | 325 |
| HLA-A*11:01 | NASVVNIQK | NASFVNIQK | 235 | 83 |
| HLA-A*11:01 | SGINASVVNIQK | SGINASFVNIQK | 411 | 150 |
| HLA-A*11:01 | INASVVNIQK | INASFVNIQK | 270 | 101 |
| HLA-A*03:02 | INASVVNIQK | INASFVNIQK | 840 | 335 |
| HLA-A*11:01 | ISGINASVVNIQK | ISGINASFVNIQK | 1056 | 450 |
| HLA-A*68:01 | NASVVNIQK | NASFVNIQK | 13 | 6 |
| HLA-C*16:01 | ISGINASVV | ISGINASFV | 368 | 787 |
N protein
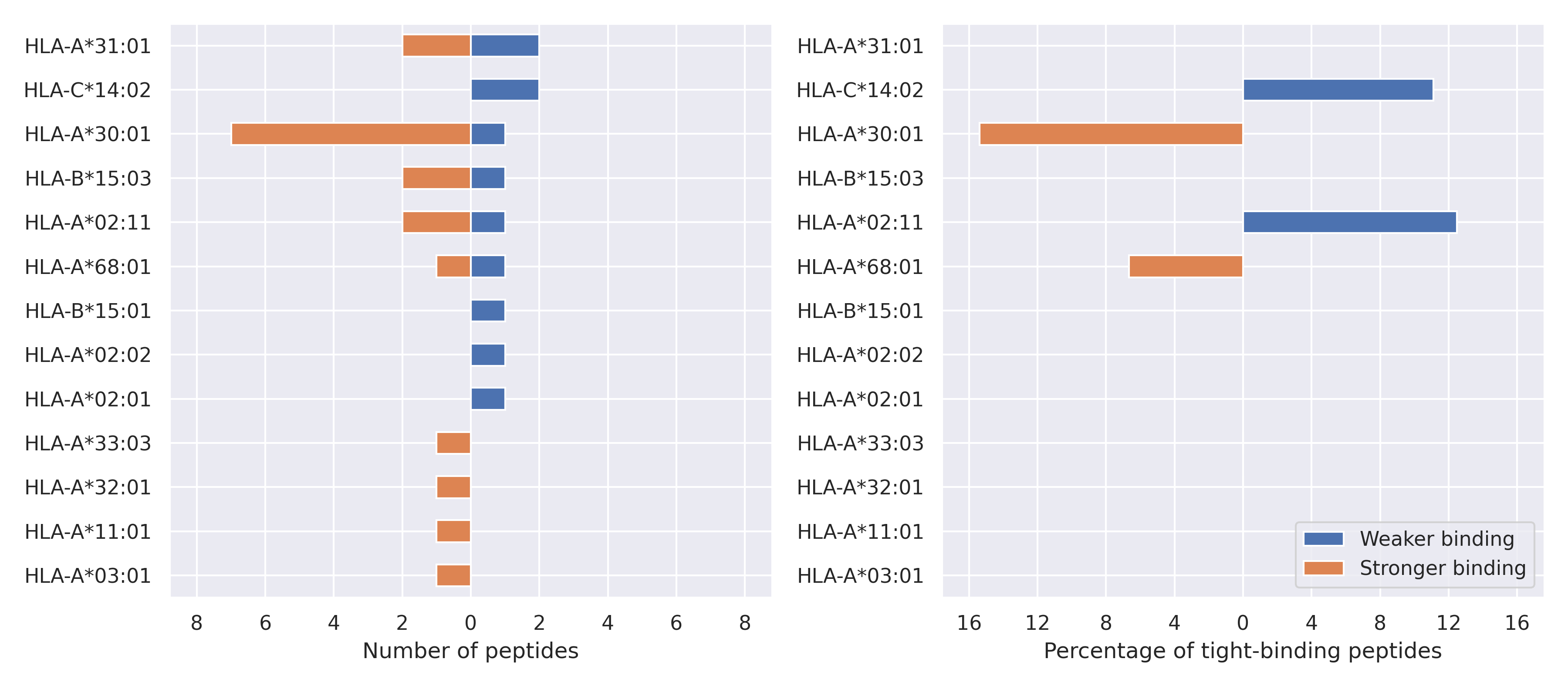
A119S
Reference IRGGDGKMKDLSPRWYFYYLGTGPEAGLPYGANKDGIIWVATEGALNTPKD
||||||||||||||||||||||||| |||||||||||||||||||||||||
Mutated IRGGDGKMKDLSPRWYFYYLGTGPESGLPYGANKDGIIWVATEGALNTPKD
| Allele | Reference peptide | Mutated peptide | Reference affinity (IC50, nM) | Mutated affinity (IC50, nM) |
|---|---|---|---|---|
| HLA-C*14:02 | YYLGTGPEA | YYLGTGPES | 40 | 275 |
| HLA-C*14:02 | FYYLGTGPEA | FYYLGTGPES | 130 | 643 |
| HLA-A*02:01 | YLGTGPEAGL | YLGTGPESGL | 218 | 576 |
| HLA-A*02:11 | YLGTGPEAGL | YLGTGPESGL | 49 | 122 |
| HLA-A*02:02 | YLGTGPEAGLPYGA | YLGTGPESGLPYGA | 155 | 318 |
203-204 RG->KR
Reference GGSQASSRSSSRSRNSSRNSTPGSSRGTSPARMAGNGGDAALALLLLDRLNQ
||||||||||||||||||||||||| |||||||||||||||||||||||||
Mutated GGSQASSRSSSRSRNSSRNSTPGSSKRTSPARMAGNGGDAALALLLLDRLNQ
| Allele | Reference peptide | Mutated peptide | Reference affinity (IC50, nM) | Mutated affinity (IC50, nM) |
|---|---|---|---|---|
| HLA-A*68:01 | NSTPGSSRG | NSTPGSSKR | 32767 | 34 |
| HLA-A*33:03 | NSTPGSSRG | NSTPGSSKR | 32767 | 300 |
| HLA-B*15:03 | SRGTSPARM | SKRTSPARM | 2669 | 106 |
| HLA-A*31:01 | RNSTPGSSRG | RNSTPGSSKR | 7013 | 340 |
| HLA-A*30:01 | SSRNSTPGSSR | SSRNSTPGSSK | 356 | 20 |
| HLA-A*30:01 | GTSPARMAG | RTSPARMAG | 493 | 30 |
| HLA-A*31:01 | SSRNSTPGSSRG | SSRNSTPGSSKR | 3757 | 231 |
| HLA-A*30:01 | GTSPARMAGNGG | RTSPARMAGNGG | 6051 | 411 |
| HLA-A*31:01 | SSRNSTPGSSR | SSRNSTPGSSK | 127 | 1853 |
| HLA-A*30:01 | NSSRNSTPGSSR | NSSRNSTPGSSK | 4255 | 353 |
| HLA-A*30:01 | RNSTPGSSR | RNSTPGSSK | 2120 | 188 |
| HLA-A*31:01 | RNSTPGSSR | RNSTPGSSK | 123 | 1335 |
| HLA-A*03:01 | SSRNSTPGSSR | SSRNSTPGSSK | 4753 | 442 |
| HLA-A*11:01 | SSRNSTPGSSR | SSRNSTPGSSK | 2231 | 263 |
| HLA-A*68:01 | NSSRNSTPGSSR | NSSRNSTPGSSK | 270 | 1385 |
| HLA-A*30:01 | GSSRGTSPA | GSSKRTSPA | 555 | 145 |
| HLA-A*30:01 | SSRNSTPGSSRG | SSRNSTPGSSKR | 1625 | 469 |
| HLA-A*30:01 | SSRGTSPARMAG | SSKRTSPARMAG | 449 | 1031 |
M234I
Reference RMAGNGGDAALALLLLDRLNQLESKMSGKGQQQQGQTVTKKSAAEASKKPR
||||||||||||||||||||||||| |||||||||||||||||||||||||
Mutated RMAGNGGDAALALLLLDRLNQLESKISGKCQQQQGQTVTKKSAAEASKKPR
| Allele | Reference peptide | Mutated peptide | Reference affinity (IC50, nM) | Mutated affinity (IC50, nM) |
|---|---|---|---|---|
| HLA-B*15:03 | RLNQLESKM | RLNQLESKI | 210 | 3572 |
| HLA-B*15:01 | RLNQLESKM | RLNQLESKI | 434 | 5192 |
| HLA-A*02:11 | LLLDRLNQLESKM | LLLDRLNQLESKI | 1721 | 220 |
| HLA-A*32:01 | RLNQLESKM | RLNQLESKI | 1285 | 261 |
| HLA-A*02:11 | RLNQLESKM | RLNQLESKI | 584 | 173 |
G238C
Reference NGGDAALALLLLDRLNQLESKMSGKGQQQQGQTVTKKSAAEASKKPRQKRT
||||||||||||||||||||||||| |||||||||||||||||||||||||
Mutated NGGDAALALLLLDRLNQLESKISGKCQQQQGQTVTKKSAAEASKKPRQKRT
| Allele | Reference peptide | Mutated peptide | Reference affinity (IC50, nM) | Mutated affinity (IC50, nM) |
|---|---|---|---|---|
| HLA-B*15:03 | GQQQQGQTV | CQQQQGQTV | 624 | 278 |
NS9c protein
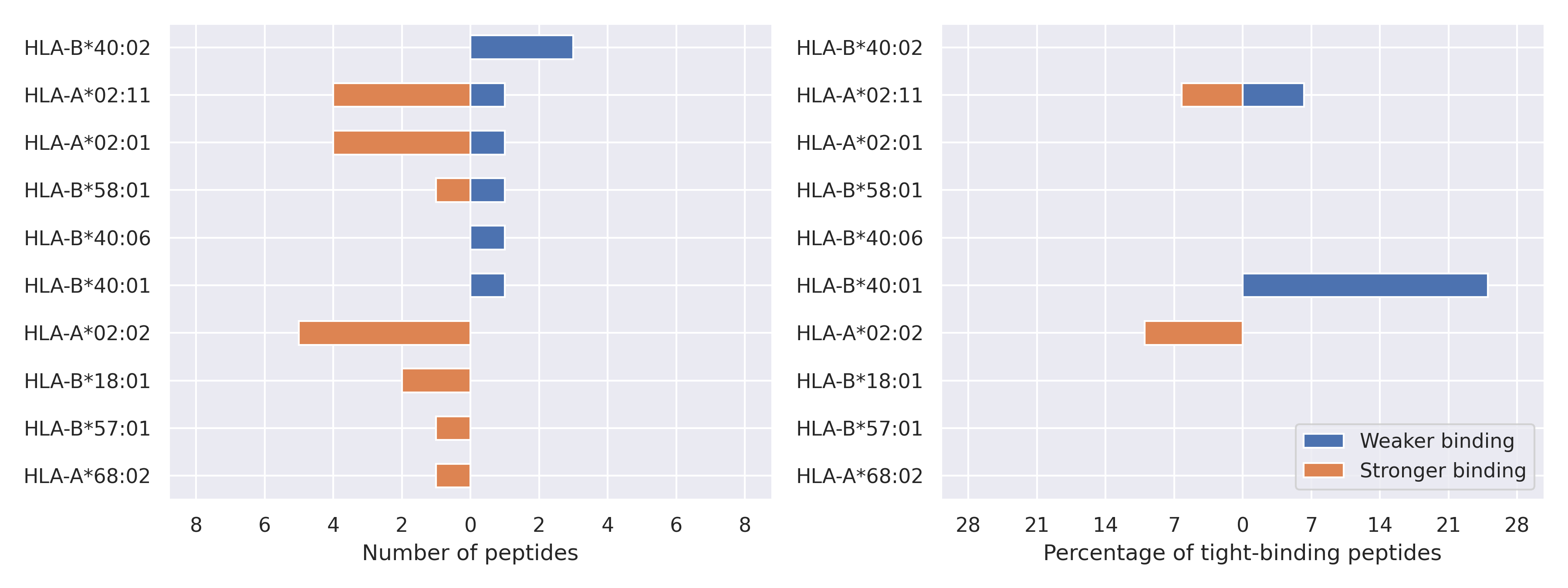
G50N
Reference AAVKPLLVPHHVVATVQEIQLQAAVGELLLLEWLAMAVMLLLLCCCLTD
||||||||||||||||||||||||| |||||||||||||||||||||||
Mutated AAVKPLLVPHHVVATVQEIQLQAAVNELLLLEWLAMAVMLLLLCCCLTD
| Allele | Reference peptide | Mutated peptide | Reference affinity (IC50, nM) | Mutated affinity (IC50, nM) |
|---|---|---|---|---|
| HLA-B*18:01 | GELLLLEWLAM | NELLLLEWLAM | 4979 | 247 |
| HLA-B*18:01 | GELLLLEW | NELLLLEW | 3915 | 401 |
| HLA-B*40:01 | GELLLLEWL | NELLLLEWL | 28 | 140 |
| HLA-A*02:11 | QLQAAVGEL | QLQAAVNEL | 120 | 27 |
| HLA-A*02:11 | IQLQAAVGEL | IQLQAAVNEL | 338 | 81 |
| HLA-B*40:02 | GELLLLEWL | NELLLLEWL | 52 | 204 |
| HLA-B*40:02 | GELLLLEWLAM | NELLLLEWLAM | 201 | 772 |
| HLA-A*02:01 | QLQAAVGEL | QLQAAVNEL | 1290 | 358 |
| HLA-A*02:02 | QLQAAVGEL | QLQAAVNEL | 92 | 28 |
| HLA-B*58:01 | VGELLLLEW | VNELLLLEW | 330 | 1071 |
| HLA-A*02:02 | IQLQAAVGEL | IQLQAAVNEL | 170 | 58 |
| HLA-A*02:01 | LQAAVGELL | LQAAVNELL | 938 | 352 |
| HLA-A*02:02 | LQAAVGELL | LQAAVNELL | 44 | 17 |
| HLA-B*40:06 | GELLLLEWLA | NELLLLEWLA | 268 | 677 |
| HLA-B*40:02 | GELLLLEWLA | NELLLLEWLA | 475 | 1195 |
| HLA-A*02:11 | LQAAVGELL | LQAAVNELL | 386 | 154 |
| HLA-A*02:01 | IQLQAAVGEL | IQLQAAVNEL | 1138 | 475 |
| HLA-A*02:01 | QLQAAVGELL | QLQAAVNELL | 961 | 403 |
| HLA-A*02:01 | GELLLLEWLAMA | NELLLLEWLAMA | 109 | 258 |
| HLA-A*02:02 | QLQAAVGELL | QLQAAVNELL | 35 | 15 |
| HLA-A*02:02 | IQLQAAVGELL | IQLQAAVNELL | 724 | 313 |
| HLA-A*02:11 | QLQAAVGELL | QLQAAVNELL | 303 | 131 |
| HLA-B*58:01 | AVGELLLLEW | AVNELLLLEW | 196 | 87 |
| HLA-A*02:11 | GELLLLEWLAMA | NELLLLEWLAMA | 29 | 61 |
| HLA-B*57:01 | AVGELLLLEW | AVNELLLLEW | 432 | 206 |
| HLA-A*68:02 | EIQLQAAVGELL | EIQLQAAVNELL | 697 | 346 |
NSP5 protein
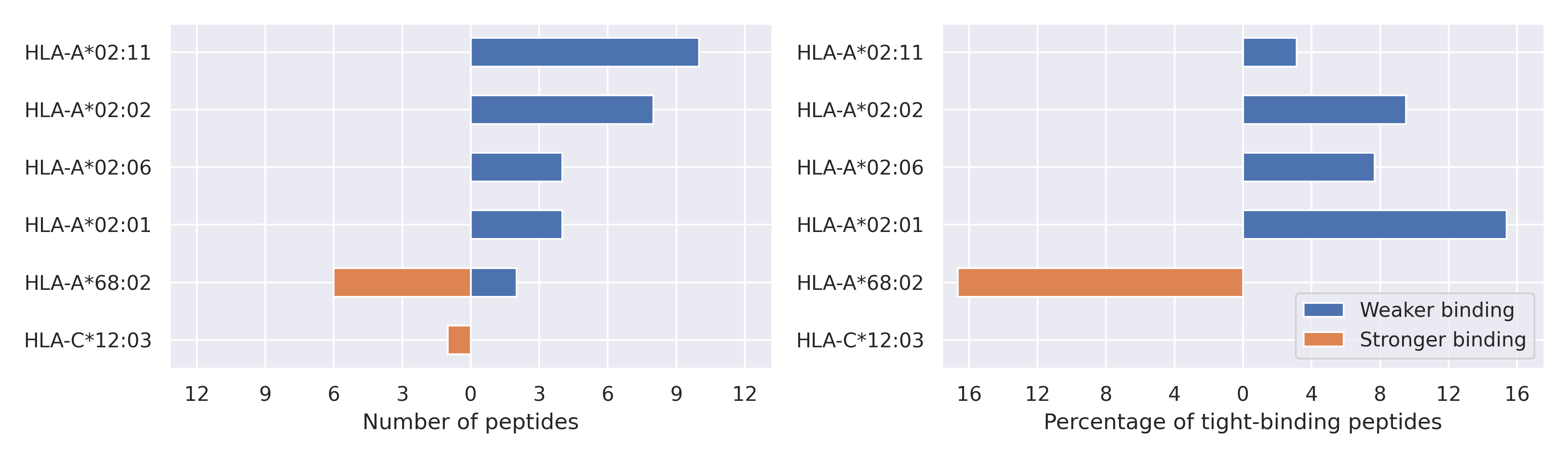
L205V
Reference NFYGPFVDRQTAQAAGTDTTITVNVLAWLYAAVINGDRWFLNRFTTTLNDF
||||||||||||||||||||||||| |||||||||||||||||||||||||
Mutated NFYGPFVDRQTAQAAGTDTTITVNVVAWLYAAVINGDRWFLNRFTTTLNDF
| Allele | Reference peptide | Mutated peptide | Reference affinity (IC50, nM) | Mutated affinity (IC50, nM) |
|---|---|---|---|---|
| HLA-A*02:11 | VLAWLYAA | VVAWLYAA | 62 | 1658 |
| HLA-A*68:02 | VLAWLYAAVI | VVAWLYAAVI | 2518 | 160 |
| HLA-A*02:11 | VLAWLYAAVI | VVAWLYAAVI | 40 | 629 |
| HLA-A*02:01 | VLAWLYAA | VVAWLYAA | 391 | 5741 |
| HLA-A*68:02 | VLAWLYAAV | VVAWLYAAV | 84 | 6 |
| HLA-A*02:02 | VLAWLYAA | VVAWLYAA | 349 | 4233 |
| HLA-A*02:11 | VNVLAWLYAAV | VNVVAWLYAAV | 80 | 828 |
| HLA-A*02:01 | VLAWLYAAVI | VVAWLYAAVI | 236 | 2140 |
| HLA-A*02:02 | VLAWLYAAVI | VVAWLYAAVI | 34 | 298 |
| HLA-A*02:11 | TVNVLAWLYAAV | TVNVVAWLYAAV | 91 | 776 |
| HLA-A*02:11 | ITVNVLAWLYAAV | ITVNVVAWLYAAV | 107 | 826 |
| HLA-A*02:02 | VNVLAWLYAAV | VNVVAWLYAAV | 192 | 1421 |
| HLA-A*02:01 | VLAWLYAAV | VVAWLYAAV | 9 | 65 |
| HLA-A*02:02 | TVNVLAWLYAAV | TVNVVAWLYAAV | 114 | 744 |
| HLA-A*02:11 | TITVNVLAWLYAAV | TITVNVVAWLYAAV | 422 | 2520 |
| HLA-A*02:02 | ITVNVLAWLYAAV | ITVNVVAWLYAAV | 152 | 761 |
| HLA-C*12:03 | VLAWLYAAV | VVAWLYAAV | 1512 | 313 |
| HLA-A*02:02 | TITVNVLAWLYAAV | TITVNVVAWLYAAV | 403 | 1845 |
| HLA-A*02:02 | VLAWLYAAV | VVAWLYAAV | 3 | 13 |
| HLA-A*68:02 | TDTTITVNVL | TDTTITVNVV | 1763 | 437 |
| HLA-A*68:02 | TITVNVLAWLYAAV | TITVNVVAWLYAAV | 1137 | 284 |
| HLA-A*02:11 | VLAWLYAAV | VVAWLYAAV | 2 | 8 |
| HLA-A*68:02 | DTTITVNVL | DTTITVNVV | 518 | 133 |
| HLA-A*68:02 | NVLAWLYAA | NVVAWLYAA | 38 | 10 |
| HLA-A*02:11 | NVLAWLYAAVI | NVVAWLYAAVI | 181 | 651 |
| HLA-A*02:02 | NVLAWLYAAV | NVVAWLYAAV | 19 | 64 |
| HLA-A*02:06 | TITVNVLAWL | TITVNVVAWL | 271 | 788 |
| HLA-A*68:02 | DTTITVNVLA | DTTITVNVVA | 65 | 182 |
| HLA-A*68:02 | TTITVNVLA | TTITVNVVA | 9 | 24 |
| HLA-A*02:01 | NVLAWLYAAV | NVVAWLYAAV | 40 | 102 |
| HLA-A*02:06 | ITVNVLAWLYAAV | ITVNVVAWLYAAV | 251 | 618 |
| HLA-A*02:11 | VNVLAWLYAA | VNVVAWLYAA | 107 | 262 |
| HLA-A*02:11 | NVLAWLYAAV | NVVAWLYAAV | 7 | 17 |
| HLA-A*02:06 | TTITVNVLA | TTITVNVVA | 483 | 1038 |
| HLA-A*02:06 | ITVNVLAWL | ITVNVVAWL | 31 | 64 |
NSP7 protein
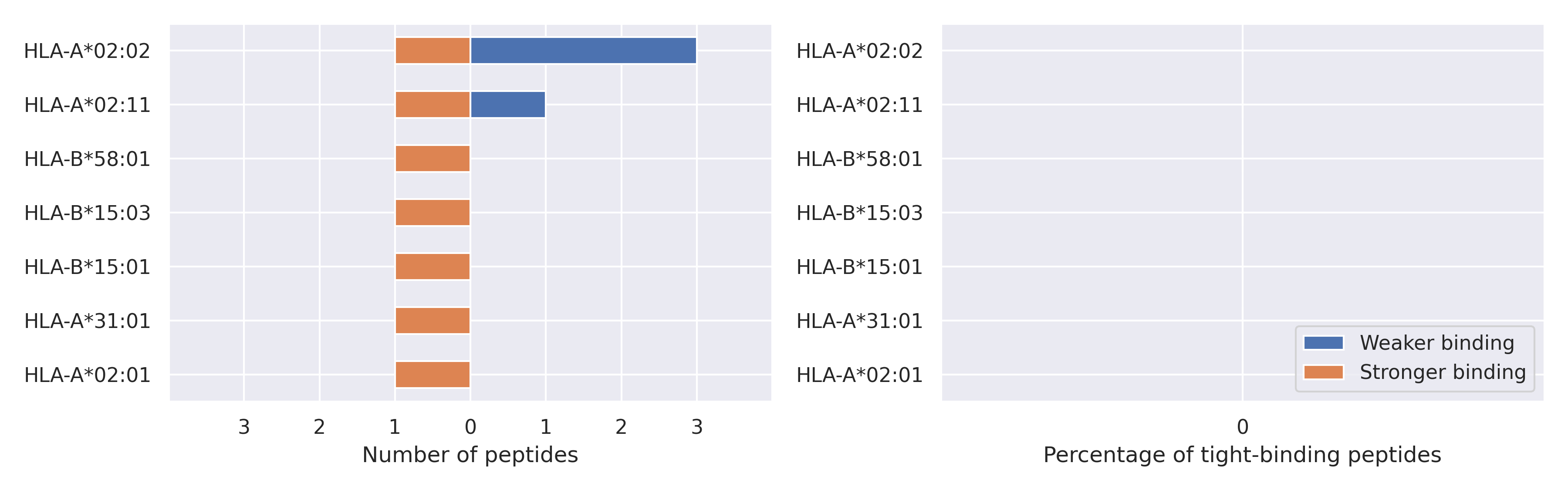
L71F
Reference TEAFEKMVSLLSVLLSMQGAVDINKLCEEMLDNRATLQ
||||||||||||||||||||||||| ||||||||||||
Mutated TEAFEKMVSLLSVLLSMQGAVDINKFCEEMLDNRATLQ
| Allele | Reference peptide | Mutated peptide | Reference affinity (IC50, nM) | Mutated affinity (IC50, nM) |
|---|---|---|---|---|
| HLA-A*02:02 | SMQGAVDINKL | SMQGAVDINKF | 382 | 10562 |
| HLA-B*15:03 | MQGAVDINKL | MQGAVDINKF | 1284 | 60 |
| HLA-A*02:02 | MQGAVDINKL | MQGAVDINKF | 359 | 7270 |
| HLA-B*15:01 | MQGAVDINKL | MQGAVDINKF | 1395 | 73 |
| HLA-A*02:11 | LCEEMLDNRATL | FCEEMLDNRATL | 709 | 64 |
| HLA-B*58:01 | LSMQGAVDINKL | LSMQGAVDINKF | 2789 | 298 |
| HLA-A*02:02 | LCEEMLDNRATL | FCEEMLDNRATL | 1191 | 137 |
| HLA-A*02:01 | LCEEMLDNRATL | FCEEMLDNRATL | 2490 | 324 |
| HLA-A*02:02 | KLCEEMLDNRATL | KFCEEMLDNRATL | 278 | 1870 |
| HLA-A*02:11 | KLCEEMLDNRATL | KFCEEMLDNRATL | 188 | 723 |
| HLA-A*31:01 | KLCEEMLDNR | KFCEEMLDNR | 673 | 240 |
NSP12 protein
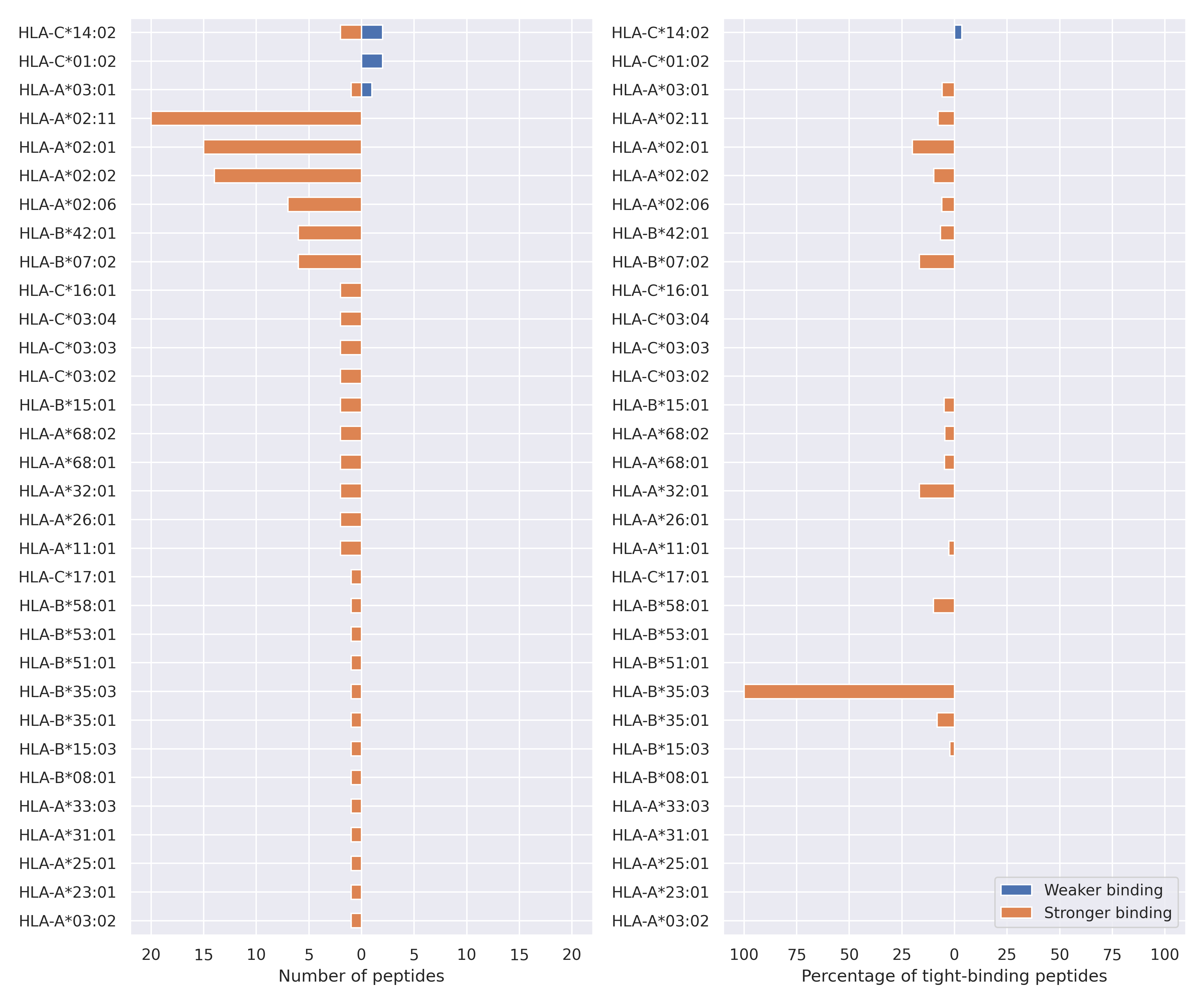
P323L
Reference CVNCLDDRCILHCANFNVLFSTVFPPTSFGPLVRKIFVDGVPFVVSTGYHF
||||||||||||||||||||||||| |||||||||||||||||||||||||
Mutated CVNCLDDRCILHCANFNVLFSTVFPLTSFGPLVRKIFVDGVPFVVSTGYHF
| Allele | Reference peptide | Mutated peptide | Reference affinity (IC50, nM) | Mutated affinity (IC50, nM) |
|---|---|---|---|---|
| HLA-A*02:01 | FPPTSFGPLV | FPLTSFGPLV | 8999 | 21 |
| HLA-A*02:02 | FPPTSFGPLV | FPLTSFGPLV | 7237 | 18 |
| HLA-A*02:11 | FPPTSFGPLV | FPLTSFGPLV | 3405 | 9 |
| HLA-A*32:01 | VLFSTVFPP | VLFSTVFPL | 5415 | 27 |
| HLA-A*02:11 | LFSTVFPP | LFSTVFPL | 6706 | 35 |
| HLA-A*02:11 | ANFNVLFSTVFPP | ANFNVLFSTVFPL | 1827 | 12 |
| HLA-A*02:11 | NFNVLFSTVFPP | NFNVLFSTVFPL | 1779 | 13 |
| HLA-A*02:11 | NVLFSTVFPP | NVLFSTVFPL | 739 | 6 |
| HLA-C*03:03 | VLFSTVFPP | VLFSTVFPL | 28468 | 265 |
| HLA-C*03:04 | VLFSTVFPP | VLFSTVFPL | 28468 | 265 |
| HLA-A*02:11 | FNVLFSTVFPP | FNVLFSTVFPL | 413 | 4 |
| HLA-C*14:02 | VLFSTVFPP | VLFSTVFPL | 21407 | 220 |
| HLA-A*02:11 | FNVLFSTVFPPT | FNVLFSTVFPLT | 2618 | 27 |
| HLA-A*02:01 | LFSTVFPP | LFSTVFPL | 7475 | 81 |
| HLA-C*14:02 | LFSTVFPP | LFSTVFPL | 11979 | 138 |
| HLA-B*15:03 | VLFSTVFPP | VLFSTVFPL | 3115 | 36 |
| HLA-A*02:11 | VLFSTVFPP | VLFSTVFPL | 78 | 1 |
| HLA-C*03:02 | VLFSTVFPP | VLFSTVFPL | 20533 | 273 |
| HLA-A*02:01 | FNVLFSTVFPP | FNVLFSTVFPL | 656 | 9 |
| HLA-A*02:01 | ANFNVLFSTVFPP | ANFNVLFSTVFPL | 2569 | 37 |
| HLA-A*02:11 | FNVLFSTVFPPTS | FNVLFSTVFPLTS | 9448 | 142 |
| HLA-A*02:02 | NVLFSTVFPP | NVLFSTVFPL | 1088 | 17 |
| HLA-A*02:11 | CANFNVLFSTVFPP | CANFNVLFSTVFPL | 6143 | 96 |
| HLA-A*02:02 | FNVLFSTVFPP | FNVLFSTVFPL | 1014 | 16 |
| HLA-A*68:02 | NVLFSTVFPP | NVLFSTVFPL | 2213 | 37 |
| HLA-A*02:02 | ANFNVLFSTVFPP | ANFNVLFSTVFPL | 3126 | 53 |
| HLA-A*02:11 | PPTSFGPLV | PLTSFGPLV | 28329 | 483 |
| HLA-C*16:01 | VLFSTVFPP | VLFSTVFPL | 24733 | 432 |
| HLA-C*17:01 | VLFSTVFPP | VLFSTVFPL | 25087 | 439 |
| HLA-A*02:01 | FNVLFSTVFPPT | FNVLFSTVFPLT | 3765 | 66 |
| HLA-A*02:02 | LFSTVFPP | LFSTVFPL | 9311 | 167 |
| HLA-A*02:01 | NVLFSTVFPP | NVLFSTVFPL | 928 | 17 |
| HLA-A*02:02 | NFNVLFSTVFPP | NFNVLFSTVFPL | 2542 | 47 |
| HLA-A*02:02 | VLFSTVFPP | VLFSTVFPL | 269 | 5 |
| HLA-A*02:01 | NFNVLFSTVFPP | NFNVLFSTVFPL | 2286 | 44 |
| HLA-A*02:06 | FPPTSFGPLV | FPLTSFGPLV | 3379 | 73 |
| HLA-A*02:01 | VLFSTVFPP | VLFSTVFPL | 179 | 4 |
| HLA-A*02:06 | FNVLFSTVFPP | FNVLFSTVFPL | 1609 | 37 |
| HLA-A*02:02 | FNVLFSTVFPPT | FNVLFSTVFPLT | 4116 | 110 |
| HLA-A*02:11 | FSTVFPPTSFGPLV | FSTVFPLTSFGPLV | 6695 | 180 |
| HLA-A*02:01 | FNVLFSTVFPPTS | FNVLFSTVFPLTS | 11451 | 311 |
| HLA-A*02:06 | VLFSTVFPP | VLFSTVFPL | 368 | 10 |
| HLA-A*02:11 | ANFNVLFSTVFPPT | ANFNVLFSTVFPLT | 14639 | 409 |
| HLA-A*02:01 | FSTVFPPTSFGPLV | FSTVFPLTSFGPLV | 12596 | 425 |
| HLA-A*68:01 | PTSFGPLVR | LTSFGPLVR | 908 | 36 |
| HLA-A*02:06 | NVLFSTVFPP | NVLFSTVFPL | 1611 | 66 |
| HLA-B*15:01 | VLFSTVFPP | VLFSTVFPL | 5731 | 243 |
| HLA-B*35:01 | FPPTSFGPL | FPLTSFGPL | 304 | 13 |
| HLA-B*53:01 | FPPTSFGPL | FPLTSFGPL | 3581 | 154 |
| HLA-C*01:02 | FPPTSFGPL | FPLTSFGPL | 217 | 4855 |
| HLA-A*02:01 | CANFNVLFSTVFPP | CANFNVLFSTVFPL | 8519 | 384 |
| HLA-A*02:02 | FNVLFSTVFPPTS | FNVLFSTVFPLTS | 10920 | 497 |
| HLA-A*02:02 | FSTVFPPTSFGPLV | FSTVFPLTSFGPLV | 6958 | 322 |
| HLA-A*02:11 | VFPPTSFGPLV | VFPLTSFGPLV | 4242 | 199 |
| HLA-A*02:02 | CANFNVLFSTVFPP | CANFNVLFSTVFPL | 8183 | 391 |
| HLA-A*02:11 | NFNVLFSTVFPPT | NFNVLFSTVFPLT | 9111 | 472 |
| HLA-A*02:06 | ANFNVLFSTVFPP | ANFNVLFSTVFPL | 5191 | 304 |
| HLA-A*02:06 | NFNVLFSTVFPP | NFNVLFSTVFPL | 4220 | 274 |
| HLA-A*02:06 | FNVLFSTVFPPT | FNVLFSTVFPLT | 4868 | 388 |
| HLA-B*35:03 | FPPTSFGPL | FPLTSFGPL | 286 | 24 |
| HLA-A*03:01 | PTSFGPLVRK | LTSFGPLVRK | 554 | 47 |
| HLA-A*31:01 | PTSFGPLVR | LTSFGPLVR | 4402 | 390 |
| HLA-A*11:01 | PTSFGPLVR | LTSFGPLVR | 4068 | 379 |
| HLA-A*03:02 | PTSFGPLVRK | LTSFGPLVRK | 868 | 88 |
| HLA-B*07:02 | STVFPPTSFGPL | STVFPLTSFGPL | 777 | 79 |
| HLA-A*68:01 | PTSFGPLVRK | LTSFGPLVRK | 308 | 34 |
| HLA-B*07:02 | TVFPPTSFGPL | TVFPLTSFGPL | 1498 | 171 |
| HLA-A*02:11 | STVFPPTSFGPLV | STVFPLTSFGPLV | 2427 | 285 |
| HLA-A*02:11 | NVLFSTVFPPT | NVLFSTVFPLT | 1183 | 139 |
| HLA-A*33:03 | PTSFGPLVR | LTSFGPLVR | 3228 | 381 |
| HLA-A*26:01 | STVFPPTSF | STVFPLTSF | 786 | 95 |
| HLA-B*07:02 | VFPPTSFGPL | VFPLTSFGPL | 339 | 43 |
| HLA-A*02:01 | NVLFSTVFPPT | NVLFSTVFPLT | 2111 | 290 |
| HLA-B*42:01 | FSTVFPPTSFGPL | FSTVFPLTSFGPL | 1320 | 185 |
| HLA-B*42:01 | STVFPPTSFGPL | STVFPLTSFGPL | 738 | 111 |
| HLA-A*02:01 | VLFSTVFPPTSF | VLFSTVFPLTSF | 2225 | 342 |
| HLA-B*08:01 | FPPTSFGPL | FPLTSFGPL | 973 | 153 |
| HLA-B*42:01 | TVFPPTSFGPL | TVFPLTSFGPL | 843 | 142 |
| HLA-A*11:01 | PTSFGPLVRK | LTSFGPLVRK | 123 | 21 |
| HLA-B*42:01 | VFPPTSFGPL | VFPLTSFGPL | 274 | 47 |
| HLA-A*02:01 | VLFSTVFPPTS | VLFSTVFPLTS | 708 | 124 |
| HLA-B*07:02 | FSTVFPPTSFGPL | FSTVFPLTSFGPL | 2537 | 466 |
| HLA-B*07:02 | FPPTSFGPL | FPLTSFGPL | 42 | 8 |
| HLA-A*02:11 | VLFSTVFPPTSF | VLFSTVFPLTSF | 1019 | 195 |
| HLA-A*25:01 | STVFPPTSF | STVFPLTSF | 1533 | 296 |
| HLA-B*07:02 | FPPTSFGPLV | FPLTSFGPLV | 1131 | 236 |
| HLA-B*42:01 | FPPTSFGPL | FPLTSFGPL | 21 | 5 |
| HLA-A*26:01 | FSTVFPPTSF | FSTVFPLTSF | 1667 | 398 |
| HLA-B*51:01 | FPPTSFGPL | FPLTSFGPL | 1809 | 433 |
| HLA-A*02:11 | VLFSTVFPPTS | VLFSTVFPLTS | 309 | 74 |
| HLA-B*42:01 | FPPTSFGPLV | FPLTSFGPLV | 368 | 89 |
| HLA-A*02:02 | VLFSTVFPPTS | VLFSTVFPLTS | 1210 | 377 |
| HLA-C*03:02 | FPPTSFGPL | FPLTSFGPL | 960 | 313 |
| HLA-A*32:01 | STVFPPTSF | STVFPLTSF | 230 | 86 |
| HLA-B*58:01 | FSTVFPPTSF | FSTVFPLTSF | 88 | 33 |
| HLA-B*15:01 | STVFPPTSF | STVFPLTSF | 130 | 49 |
| HLA-A*02:01 | VLFSTVFPPT | VLFSTVFPLT | 61 | 23 |
| HLA-C*03:03 | FPPTSFGPL | FPLTSFGPL | 375 | 145 |
| HLA-C*03:04 | FPPTSFGPL | FPLTSFGPL | 375 | 145 |
| HLA-A*02:02 | VLFSTVFPPT | VLFSTVFPLT | 113 | 44 |
| HLA-C*16:01 | FSTVFPPTS | FSTVFPLTS | 1127 | 460 |
| HLA-C*14:02 | LFSTVFPPTSF | LFSTVFPLTSF | 396 | 950 |
| HLA-C*01:02 | VFPPTSFGPL | VFPLTSFGPL | 429 | 997 |
| HLA-A*02:02 | TVFPPTSFGPLV | TVFPLTSFGPLV | 789 | 343 |
| HLA-A*68:02 | STVFPPTSFGPLV | STVFPLTSFGPLV | 169 | 74 |
| HLA-A*02:11 | VLFSTVFPPT | VLFSTVFPLT | 25 | 11 |
| HLA-A*03:01 | TVFPPTSFGPLVRK | TVFPLTSFGPLVRK | 92 | 198 |
| HLA-C*14:02 | VFPPTSFGPL | VFPLTSFGPL | 47 | 99 |
| HLA-A*23:01 | LFSTVFPPTSF | LFSTVFPLTSF | 370 | 179 |